
good tribe / bad tribe — nerdcore mix
Theories of Human Evolution
Reading Dawkins
Dawkins’ Selfish Gene carries a lot of weight.
I discovered that back in 1975 when I first threw it across the room.
This happened right after the opening paragraph where he dismisses all philosophy written before Darwin. I hadn’t even gotten to the ‘lumbering robots’ part twenty pages in.
That was another toss.

Like his comments on the worthlessness of all pre-Darwin philosophy in the opening paragraph, his compelling descriptions of George Jensen’s work on gene selection…a simplicity at the heart of natural selection…is both useful, rhetorically crisp, and massively overgeneralized.
My attitude towards Dawkins was not improved seeing him cited in the business press throughout the ‘greed is good’ 80's.
This essay is about the maturing of models and a deeper, more or less terrifying, view that a these more complex models give us of human nature.
It argues for group selection: George Jensen’s (and Dawkins’) bogey man.
A quick look at some conclusions before we take a more detailed trip to get there:
- Group selection is a strong factor in human evolution. We see it daily in our deeply ingrained Us vs Them thinking.
- Groups win by beating other groups. The genetically rooted behavioral tendencies of any individual can determine whether a tribe lives or dies. This is where group selection is forged. A ‘selfish gene’ for self sacrifice can be selected at the tribal level.
- This all requires a emphatic bond to the folks we consider to be us. This ‘us’ has a flip side. Another group of our own species is ‘them’ and the ability to empathize and cooperate with them has to be blocked. A deep animosity has to take it’s place.
- In other words, ‘us’ and ‘them’ are intimately paired by human evolution. Evolution has given us the capacity for deep empathy and genocidal warfare; it has given us a smorgasbord of altruism, cheating, empathy, love, hatred, war, and genocide.
- As we’ll see, the emotional forces that advance pro-Us and anti-Them action are even intertwined by the neurotransmitter oxytocin on a molecular level. (tl/dr? Skip down and watch the Sapolsky video.)
- Group selection isn’t a fuzzy theory. It can be modeled to test suppositions and boundary conditions. This, however, was difficult to implement until more recently with the advance of computing power. I’ll trace the history.
Scientific Progress
This is in part, then, a story of how scientific models evolve from simple to complex. It is, also, a story about how even the simplest models can highlight deep structures in our identity and our personal and collective struggles.
It is important to understand how science progresses. (I’m using Thomas Kuhn’s insights here.) Opponents of Relativity and Quantum Mechanics did not generally covert from physic’s old paradigm. They were supplanted by the newer theory and it’s adherents.
Further, and critically, evidence for the new theory is gathered under the old theory as it boundaries are pushed beyond what they can successfully predict and as anomalies accumulate.
The job of scientists of any model is, therefore, to push it to the limits defining through experimental hypothesis testing what it can account for and where the model starts tripping over its own limitations. That’s progress.
Dawkins’ insights aren’t negated but have, imho, been reframed by later, more complete and complex models. Dawkins pushes the selfish gene model to the limits insisting that it, alone, is sufficient. In a sense, you can’t blame him; that’s what science does: push the limits.
Yet I do blame him.
Science should be about pushing the limits of a theory through research…not rhetorical arrogance.
Q: But for now it seems like the bulk of scientific opinion is against you? A: Science is not done by polling. Have you ever heard of “100 Scientists Against Einstein?” It was a pamphlet signed by 100 physicists to overthrow his theory of relativity. After they published it, Einstein remarked, “Why 100 authors? If I were wrong, then one would have been enough!” - E O Wilson defending the group selection model in Discover Magazine.
Cheaters always win
Scratch an ‘altruist’, and watch a ‘hypocrite’ bleed - Michael Ghiselin, (1974) The Economy of Nature and the Evolution of Sex
This is, in part, a story of how scientific models evolve from the simple to the complex. It is, also, a story about how even the simplest models can highlight deep structures in our identity and our personal and collective struggles.
In the mid 60’s George C, Williams proved that altruism couldn’t exist. It’s all a matter of math and if evolutionary strategies can be stable over the long term.
Consider the following three cases.
- Completing an action that makes one fitter ( i.e. more likely to add to the gene pool) bears a cost. You spend energy to gather food and the food keeps you alive to reproduce and keep your offspring alive.
- An altruist bears a cost while sharing the benefit. You gather food and share it with someone that doesn’t share a distinguishing genetic connection. You’ve spent the same energy but with less return to your evolutionary fitness.
- A cheater gains the benefit without bearing the cost. You head out to gather, grab a few nuts, take a nap in the shrubbery, and return claiming bad luck…but eat, nonetheless, out of the communal pot.
Slowly over the generations, the altruist strain weakens and dies out.

The key concepts here are relative genetic fitness and an ‘evolutionary stable strategy’. Fitness is defined as reproductive success. The more offspring that survive to reproduce, the more the set of underlying genes thrive.
Imagine green eyes give an advantage and blue eyes don’t. You can’t have both green eyes and blue eyes. Unless there are additional complicating factors, over the generations blue eyes will go away.
Importantly, in a population of blue eyes, a green mutation can invade and take over.
To thrive, a trait must be an evolutionary stable strategy, i.e. both robust and able to resist invasion by competing traits.
Parallel to green eyes and blues, Altruism and Selfish are competing genes; you can’t be both. In a Total Population you’d then have two subgroups: Altruists and Selfish. (Cheaters are a subset of Selfish for the discussion below.)
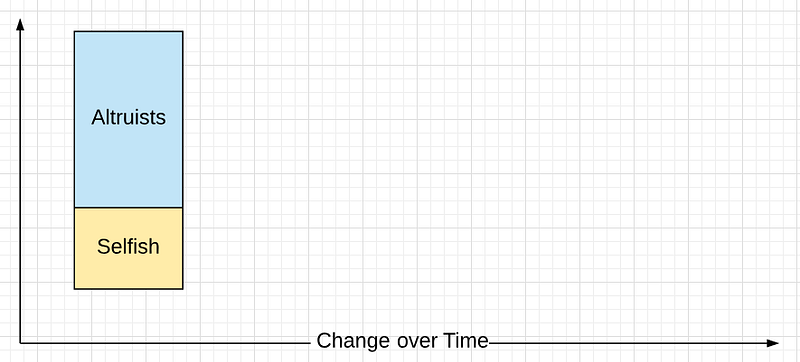
In each generations, the two groups increase or decrease in population frequency within the Total Population.
The Size of Selfish is adjusted by (Base Fitness plus Altruist Benefit).
The Size of Altruists is adjusted by (Base Fitness plus Altruist Benefit) minus Cost of Altruism.
(Base Fitness plus Altruist Benefit) is necessarily greater than (Base Fitness plus Altruist Benefit) minus Cost of Altruism) just as (2+2 ) > (2+2) -1.
Why? Benefits are distributed across the whole population but the cost is born by a few. The Size of Selfish in each succeeding generation is necessarily greater than the Size of Altruist since selfish has no cost and that’s always cheaper than the Cost of Altruism. Cost and benefits, to repeat, are expressed in impact on fitness, i.e. reproductive success.
Given enough generations, Selfish drives Altruists to extinction.
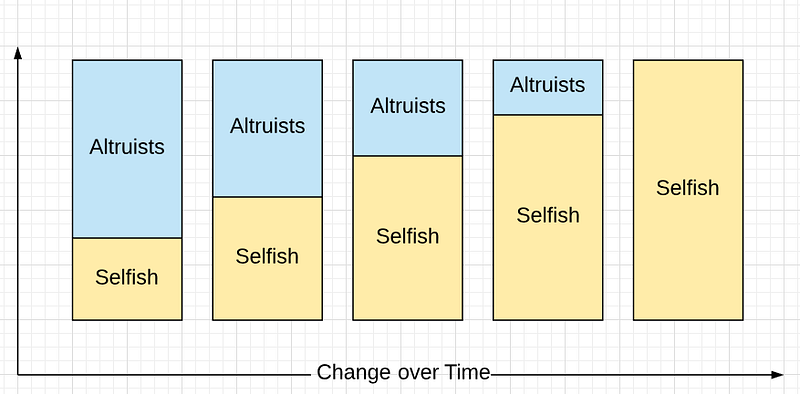
In the genetic arms race, cheaters always win.
Or do they?
That seemed pretty simple. Too simple.
I’m going to de-simplify in two steps. The first step involves a phenomena called Simpson’s Paradox. The second involves the human propensity to war and genocide.

Step One — Haystacks
Imagine a field full of haystacks. (This follows a classic thought experiment from one of the key theorists in evolutionary biology, John Maynard Smith.)
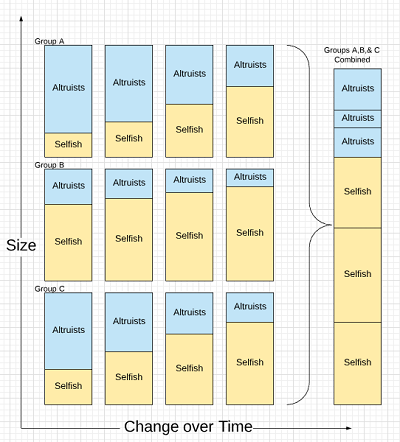
- When the haystacks are built they each have a random mix of Selfish and Altruist mice assigned from the total population.
- The mice breed through the winter for X number of generations until the haystacks are consumed and a new field planted.
- The population is thereby recombined until the haystacks are built again the following fall.
Step one, therefore, starts with multiple populations separated by space (the haystacks). Each group remains separate for some time period but then recombines.
Well, nothing changed. The size of Altruists is always bigger at the start than at the end. Selfish thrives; Altruists decline. The sum of extinctions over time is still extinction.
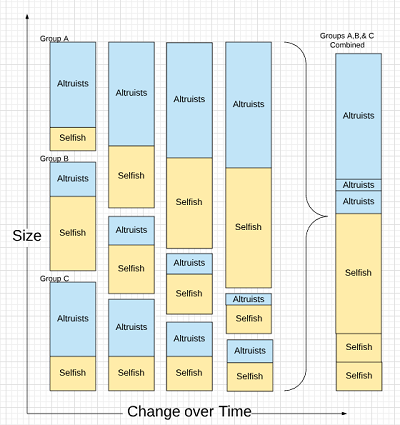
But size is misnomer. What’s really being shown is frequency. The percentage of Selfish is what increases in each group. But if we consider that Altruists might be an advantage to the group, then the size of the populations with more Altruists will increase faster than the size of a population with fewer.
Another way to say this is that a group with lots of Selfish or, worse, Cheaters isn’t going to do that well.
This is Simpson’s paradox: despite an increase of Selfish in each group, the total number of Altruists can remain stable overall as the groups with a strong Altruist population thrive.
The images above are a sloppy way to try and prove a point…and they aren’t intended to. The objective is to communicate the general concept.
To go further would require switch to a mathematical version of the model. Components, (the ratio of Altruists to Selfish in each generation, the cost and population growth benefits of altruism, etc.) need to be expressed numerically so that border conditions, stable strategies, and tipping points can be discovered. Clearly, if the cost of Altruism is too high or the benefit to the group too low, the possibility raised above is moot.
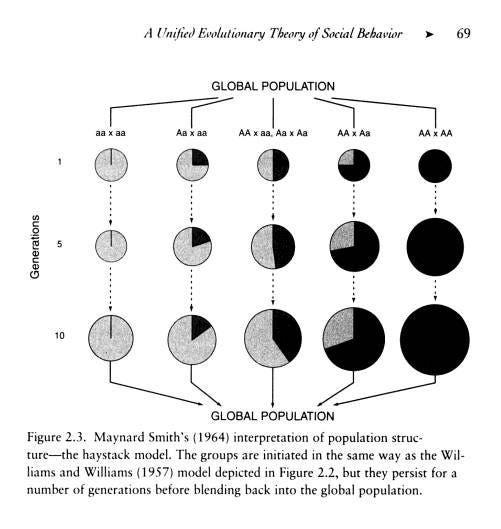
Here’s an example of David Sloan Wilson working out on a mathematical model. (And, just for shits and giggles here’s my 2 cents on mathematical models.)

How does human evolution match the haystack model?
We’ll not so good. We evolved in pretty leaky haystacks, for example.
The model does works but in some odd situations, e.g. parasites and slime molds, where you get clear genetically isolated ‘haystacks’ and then a remixing.
As to humans, consensus was that you can’t get there from here, all things being equal…but, of course, all things aren’t…
War is the answer

Perhaps you can’t get there from here but nonetheless there clearly is a there there.
We need to account for something rare.
Human beings are an outlier on a scale of ‘prosociality’…an outlier among all mammals, all primates, or even among our own chimpanzee wing of the ‘higher apes.’ An individual human, or “me, my old lady, and my dog” have a near zero survival value. Our evolutionary success clearly required tribes and we take tribes to a level without any real parallel even among highly social species. We’re somewhere up there with ants, parasites, and mole rats in sociality despite lacking some of the crisp genetic transmission uniquenesses that underlie the social structure of all but us on the list.
Human behavior provides a wide variety of examples of behavior that require a cost and that benefit someone in addition to the actor…clearly in cooperation but, also, in apparent altruism. How do we get there?
(Why talk about the haystack model at all, then? Models are ways to clarify our thinking; the model of competing groups with rules governing gene exchange between groups is a great place to start.)
In Step One above I described what is essentially a foot race. Each group is in its own lane, as it were. It isn’t competing for resources. There’s no interaction until the race is over. The winner is the group with the highest population at the end. They end up being disproportionally represented in the mixer party at the race’s end.
Step Two — Conflict
For Step Two, it’s a more directly competitive game. Your team can get out of its ‘lane’ and mess with your neighbors. You can directly compete for resources and the more successful competitors can starve out their neighbors.
Worse, you can cross the lane lines and conspire to kill members of other groups.
Human tribes do that.
Further, the only other mammal species that conducts genocidal raids on conspecifics is one of our two closest relatives, the chimp species, Pan Troglodyte.
Offsetting the fuzzy boundaries of our ‘haystack’ in the genetic arms race is our ability to strike out and set fire to other haystacks.
The modeling, now, gets more complex than the simple formulas that drive the previous discussion; simple formulas can open the door to possibilities but they can’t really work with the tipping points and trade-offs with sufficient precision to meaningfully test this hypothesis.
With recent computing power, however, it is possible to model out hundreds or even thousands of individual actors as agents and set ‘personal’ variables for sacrifice, cheating, retaliation against cheaters, and behavior in inter-group conflict. It’s then possible to set up test scenarios and see what the agents do in repeated runs.

For example:
“Using a model of the evolutionary impact of between-group competition and a new data set that combines archaeological evidence on causes of death during the Late Pleistocene and early Holocene with ethnographic and historical reports on hunter-gatherer populations, I find that the estimated level of mortality in intergroup conflicts would have had substantial effects, allowing the proliferation of group-beneficial behaviors that were quite costly to the individual altruist.” - Sam Bowles, Did warfare among ancestral hunter-gatherers affect the evolution of human social behaviors?, Science June 5, 2009
Human haystacks may be more permeable but there’s an additional lethal dynamic at play that counter-balances that with a very direct form of natural selection that works specifically at the group level: the groups that are the tightest, that are the most effective in group action, will be the more successful in attack or the more resistant to attack through coordinated defense. Individual self-sacrifice might save a group at the cost of an individual.
Cheaters in this context are called traitors and are typically summarily executed.
In the terms I’ve adopted, the strongest ‘us’ wins out. Groups win by beating other groups and the genetic tendencies of any individual can determine whether a tribe lives or dies en masse.
This ‘us’ has a flip side. Another group of our own species is ‘them’ and the ability to empathize, connect, and cooperate with the ‘other’ people has to be blocked and a deep animosity to ‘them’ has to take it’s place.
Genocidal raids are rare among species. So is our high level of cooperative empathy. The simplest explanation is that they co-evolved from a single root.
War! A Short History
(The evolutionary path I’ve sketch above is reliant on “group selection” rather than kin selection / inclusive fitness and highly controversial. If you want to dig into that a bit, I’ve a discussion here. It worth checking out only if you want to get seriously into the weeds.)
I think it’s important to get a sense of what war means “on the ground.” Below, we’ll look at the neurochemistry and emotion driving the us/them split. First, an outside in look at the progression or genocidal raids from it’s likely prehuman origins to recorded history.
If you’re uninterested in this, fairly lengthy, chronicle, skip to the Truth Is Within section below.
1) Chimps tribes conduct genocidal raids on other chimp tribes
Chimps are extremely rare among species in that groups of chimps make genocidal attacks on other against groups of chimps. (The argument that this is abnormal chimp behavior lacks evidence.)
2) Prehistory: there’s archaeological evidence of the same among early humans
In addition to the chimp research, there’s accumulating evidence from prehistory. For example:
“Whoever the assailants were, they probably arrived at dawn. Catching their victims unawares, they hacked the shinbones of as many villagers as possible to prevent them escaping, then bludgeoned them all to death with blows to the head before dumping them in a mass grave. Though no one can be sure exactly what happened to the 26 people whose bodies were found in a Stone-Age mass grave, this is the most likely scenario….”
3) Ancient Warfare was, typically, genocidal, too….with much higher casualty rates

I’m picking Greek hoplite warfare because there’s literally centuries of research into it.
Hoplite warfare was generally conducted on a flat plain with two lines of hoplite warriors in ranks up to 8 deep colliding with each other in a battle frenzy. The soldiers were not a special caste but citizens who provided their own gear, generally a special shield, a short sword, and a spear typically with metal points on both ends. The butt end point was for finishing off the wounded if they got trampled under foot.
A key feature of most ancient warfare was highly asymmetrical casualty rates. Once the enemy’s line broke, their retreat became a massacre. A primary use for cavalry and even chariots in ancient war was to chase down and slaughter retreating soldiers. Selfish soldiers that decided to flee and save their own skin could cause the line to crumble with disastrous results. And, as mentioned above, running away, unless part of a general panic and rout, typically led to an execution for desertion.
From a Reddit post, What was the casualty rate for battles between hoplites in ancient Greece? (The post cites a variety of source which, unfortunately, hide behind a paywall. I’ll add a link to the most relevant below.)
The delight felt by the Greeks when their enemies ran — and I do mean delight; Xenophon once refers to the pursuit as one of the ‘pleasures’ of war (Hieron 2.15) — shows that the clumsy, awkward encounter of poorly trained hoplite masses wasn’t the ultimate goal of having a battle. The goal was to break the enemy, so that they could be pursued and killed with impunity. For the same reason every Greek citizen army fought to keep its men alive, they also fought to annihilate their enemies: every fatality directly impacted the enemy community, and enough death could destabilize their whole state. There was a great deal to be gained from battle if the enemy casualty rate could be made as high as possible.
We see the effect of the pursuit clearly reflected in the death toll of hoplite battles. At the Nemea in 394 BC, the Spartans are said to have butchered 2800 of their opponents in flight; at Kynoskephalai in 364 BC, Thessalian cavalry massacred over 3000 of the men of Alexander of Pherai…. Such losses would have cut swathes through the citizen body of the states involved. It is not surprising that Argos, 15 years after the defeat at Sepeia, pleaded that it had no choice but to stay neutral in the war against Persia, because it had no grown men.
The Truth is Within
Things are the way they are because they got that way. - Gerald Weinberg
Mathematical models are inherently ‘bloodless’...abstract formulations that help define boundary conditions, trade-offs, and interconnections. What I find fascinating is how clearly the models described above point to the internal conflicts of our species…to the shadings of our emotions, impulses, and cognitive frameworks. The ‘bloodless’ models foot out to altruism, cheating, empathy, love, hatred, war, and genocide
Watch out for the Cheater
First, if a haystack-like dynamic pertains, then one would expect an ongoing battle between our tendency to cheat and our tendency to detect and try and punish cheaters. The evidence is within.
Exhibit 1
We seem to share an baked in sense of ‘fair’ with our close relatives on the photogenic tree. In 2003, Brosnan, Wolkenten, and deWaal (a personal hero of mine) published some watershed research using Capuchin monkeys.
Franz deWaal recaps the results here:
A summary from the scientific press: “…[if] one monkey …landed a grape, while monkey number two got a cucumber, madness ensued.”
A monkey that feels cheated is one angry monkey. From there it’s a short hop to road rage should some driver feel cut off in traffic or to Ronald Reagan’s political use of the mythical ‘welfare queen’.

Exhibit 2
Humans are not all that good at logic. Even college undergraduates (the lab rats of many psychology experiments) get less than a third of the questions right on commonly used tests.
Here, for example, is a simple logic test, the Wason Selection Task test.
How many cards do you need to turn over to test the rule: if the card shows an odd number on one face, then its opposite face is brown? Only a card with both an odd number on one face and something other than brown on the other face can invalidate this rule. How do you tell if it is true of the card set below?
Try to answer before scrolling down. Less than 10% of the lab rats got it right.

Evolutionary psychologists, Cosmides and Tooby did a fascinating variant.
We’re going to test a different rule: If you are drinking alcohol then you must be over 18. The cards have an age on one side and what that person is drinking on the other. What cards do you have to turn over to find out if everyone is following the rule?

Easier right? The correct answer in both cases is cards 1 and 4. Not surprising many more folks get this variant right.
Yet those are precisely the same logical puzzle.
Daniel Dennett unpacks this in his book, Darwin’s Dangerous Idea:
“If…questions are restated to provide a social context of fairness, just entitlement, or cheating, then success doubles! These results remain robust across cultures, ages, and backgrounds. …the easy cases are all cases that are readily interpreted as tasks of patrolling a social contract, or, in other words, cheater detection.
It seems that human beings are hard wired to detect cheaters!

Cheatin’ Conclusions
1) Cheater Detection, since it has likely persisted over some millions of years, likely has a strong genetic underpinning as an important activity in the reproductive success of primates including homo sapiens.
2) That and other evidence indicates that Cheating has also persisted…whether or not it is similarly genetically encoded. There’s no need to have a built in Cheater Detection system if it has not been an ongoing problem.
3) This persistence of both would seem to provide prima facie evidence that Cheating and Cooperation enforced by Cheater Detection are what’s termed ‘evolutionary stable strategies’ with neither able to eradicate the other.
Love and War
I’ve only got one exhibit here but it’s a remarkably telling.
Moving from intra-tribe dynamics to inter-tribal conflict we do, indeed, find traits that are remarkably rare among species:
- First, we an outlier in pro-sociality…near unique among vertebrates: the depth of our connection to a group of other humans has no real parallel.
- Second, we’re an outlier in our tendency to join in with our group and try to eliminate other groups of humans.
We war on conspecifics. We sort our species into ‘us’ and ‘them’. (And in the broad stroke of human prehistory it is almost certain that our ‘them’ contained a few children of ‘us’.) It’s instinctual ‘fast-thinking’ based on threat analysis. Them’s have been dangerous!
Disconnected now from much of the physical danger, we still seem to need us vs them.
Again “the truth is within”
Exhibit 1
Conclusions first; evidence second.
- Research into the biochemical basis of bonding has focused on a hormone, oxytocin
- Paradoxically, oxytocin makes out-groups more them-like!!
- In other words, the emphatic basis of deep cooperation and the ability to view conspecifics as a dangerous and hated other are promoted by the same neurotransmitter.
- This makes sense if a strong connection to folks on my ‘team’ and a equally strong antipathy to ‘them’ arose with warfare as a driver of primate and then human evolution.
First, Love is the Drug…
It’s worth watching both the following videos and, in particular, the full Robert Sapolsky video! I’ll offer transcription excerpts to give a few key points.
The Paul Zak video is particularly rich from 6:30–8:45 in discussing oxytocin. There’s a nice bonus at 11:20 on cheater punishment.
(the link, just in case the video fails to run)
6:30–8:45 So, oxytocin is the trust molecule. Oxytocin increases generosity by 80%. What does it feel like when your brain is flooded with oxytocin? The change in oxytocin predicted feelings of empathy. It’s empathy that connects us to other people. It’s empathy that makes us help other people. It’s empathy that makes us moral.
11:20 There’s another way oxytocin is inhibited which is interesting: through the action of testosterone. In experiments we have administered testosterone to men. Instead of sharing they become selfish. But, interestingly, high testosterone males are, also, more likely to use their own money to punish others for being selfish.
… a Double-edged Drug at that
2:25 Your brain processes these us/them differences on the scale of millisecond…a 20th of a second your brain is already responding differently to an us vs a them. So collectively, this is as depressing as hell. Oh my god, we are hardwired to inevitably be horrible to thems and thems along all sorts of disturbing lines. But…
4:30 Okay, a fabulous study showing this double edged quality to oxytocin. And this was a study done by a group in the Netherlands... They gave [students] a classic philosophy problem, the runaway trolley problem, is it okay to sacrifice one person to save five. Runaway trolley, can you push one…onto the track to save five…. You give people this scenario and they have varying opinions. Now you give them the scenario where the person you push onto the track has a name. them give them names…either Dirk or names that invoke lots of xenophobic hostility…Otto or Mahmoud. In general, given those names, and there’s no difference in how people would rate it than if they were anonymous. Give people oxytocin and, kumbaya, you’re far less likely to push Dirk onto the track…and more likely to push good old Otto or good old Mahmoud onto the track. All you’ve done is exaggerate the us/them divide with that.
Voila, the hormone that makes ‘us‘ more Us, makes ‘them’ more Them.
And a key insight, Us vs Them thinking is fast thinking. We react before we think. The us/them divide is hard-wired into our brains.
Coda: Religious Genius

Can we evolve beyond a construct of constant enemies, constant crisis?…It is possible that this whole book is about love. - adrienne maree brown (2017) Emergent Strategy
But the end is reconciliation; the end is redemption; the end is the creation of the beloved community. It is this type of spirit and this type of love that can transform opposers into friends. The type of love that I stress here is not eros, a sort of esthetic or romantic love; not philia, a sort of reciprocal love between personal friends; but it is agape which is understanding goodwill for all men. It is an overflowing love which seeks nothing in return. It is the love of God working in the lives of men. This is the love that may well be the salvation of our civilization. -Martin Luther King, Jr (1957) “The Role of the Church in Facing the Nation’s Chief Moral Dilemma”
The golden way is to be friends with the world and to regard the whole human family as one. -Mahatma Gandhi (2005). “All Men Are Brothers”, p.115, A&C Black
So then, it seems that we’re naturally good…
… and we’re naturally monsters.
We can “love our neighbor as ourselves”; and we evolved under threat from other humans. We survived by assuming that there is risk and a danger of attack before waiting for the proof.
What do we do with this weird bag of parts we’re given to assemble our lives and culture?
I have a theory of ‘religious genius.’
We all have a deeply held feeling for tribal connection…a sense of ‘us’… a fellow feeling with others of our family and tribe. Evidence is clear that deprived of connection we suffer from increased mental illness and physical disease. Similarly, connection and regular interaction with our personal “beloved community” makes us more robust and happier.
And we all have a tendency to dehumanize other humans…to blame them, even persecute and war on them.
Religious genius, I believe, can see through that immediate tribal connection of ‘us’ out into a connection with a wider Us.
My guess is that the Martin Luther Kings and Gandhis of human history are people that felt the contradiction most acutely and resolved it, after a profound inner struggle, by widening out their sense of compassion to the greatest possible Us…to their mortal peril and to the advancement of us all.
What about you and I?
Well, I don’t think most of us can hope to achieve religious genius…but couldn’t we, just maybe, aim for some sort of religious sub-genius status?
Could we recognize that taking many small steps to achieve an only slightly bigger ‘us’ can have, cumulatively, a huge impact.

Using prayer or meditation to suspend our immediate negative reaction is a good start. There are more resources here.

(Errata: Oops, correction. The quote I attribute to George Clinton was originated by Charlie Wilson of the Gap Band in ‘I Don’t Believe You Want to Get Up and Dance’. I had my funk chronology reversed. Inexcusable.)
References:
Subscribe
You can sign up below. I’ll let you know when I publish something.