
Viruses reveal the secrets of plasmodesmal cell biology
Plasmodesmata (PD) (singular: plasmodesma) are highly specialized trans‒wall channels that function as gateways between cells. Plasmodesmata build connections between neighboring cells and facilitate the transport of macromolecules such as proteins, carbohydrates, ions, and RNA complexes: messenger RNA (mRNA), transfer RNA (tRNA), green fluorescent proteins, and RNA proteins between cells.
First, Plasmodesmata were discovered by Edward Tangl in 1885 and confirmed their presence in higher plants. Later, it is found that they are present in all green plants. They are characterized as communication channels that form long, cylindrical bridges that pass the cell wall of each cell and form connections.

Structure of Plasmodesmata
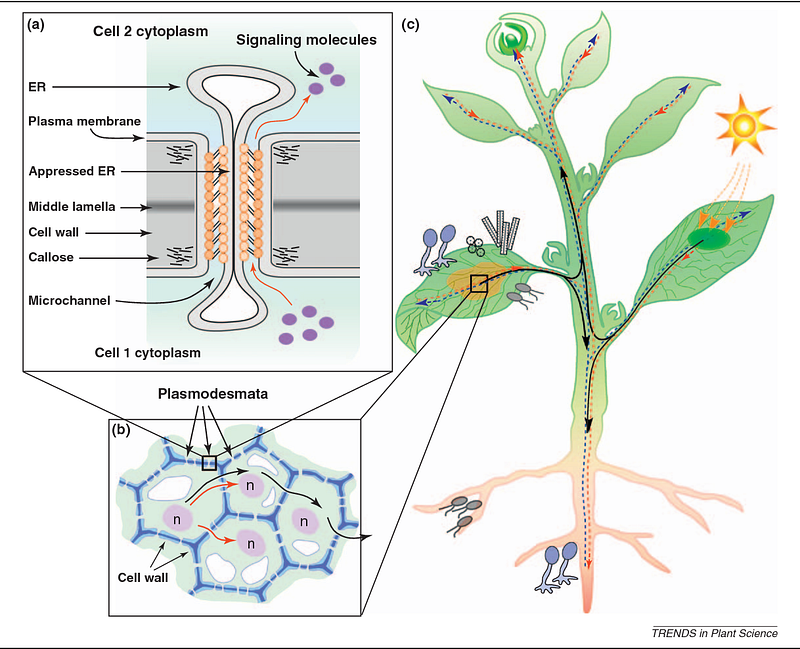
Plasmodesmata forms aqueous and cylindrical bridges that transverse the cell wall and builds an interconnected network in the whole plant.
Their diameter is approximately 40 nm. These junctions consist of three layers: the plasma membrane, the cytoplasmic sleeve, and a desmotubule. The desmotubule is a tube of the endoplasmic reticulum that runs between two adjacent cells.
Desmotubule is electron-rich particles with 2.5 nm in diameter.
Transport Pathways in Plants
Plants have evolved specialized mechanisms for the exchange of water, nutrients, photosynthates, and signal exchange. For the transport of food and water, plants have conductive tissues (xylem, phloem).
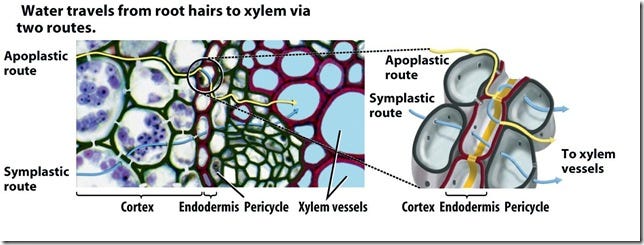
These vasculature tissues are used for long-distance transfer while for short term transfer, plants have an apoplastic/symplastic pathway. In the symplastic pathway, transport occurs through plasmodesmata: which plays a vital role in intercellular communications.
Previous studies suggest that Plasmodesmata are static (nonselective) entities that encourage all kinds of molecules but recent advances in Plasmodesmata biology proved that these are highly selective channels with the ability to “gate”.
Plasmodesmata build an interconnected cytoplasmic network throughout the plant body that supports the transport of the targeted molecules into the neighboring cells.
Plasmodesmata provide a direct cell to cell connection and also facilitate the direct transport of macromolecules without the limitations of the rigid cell wall.
Plant Pathogens Exploit the Plasmodesmata
Plasmodesmata play a crucial role in plant cell communications and plant pathogens always try to breach these channels and their regulations. Because the primary purpose of microbes (pathogens) is to infect plants and then, spread systemically.
Therefore, they exploit the trans wall channels, especially plant viruses enter into the plant’s vasculature to spread into the plant system. Surprisingly, plant pathogens found these microchannels as an opportunity to spread systemically for infection.
Fungal pathogens also exploit plasmodesmata for their spread in plant hosts. Particularly, hemibiotrophic fungi spread through plasmodesmata to cause infections in plants.
Many studies proposed that these pathogens might have the ability to modulate or remodel the plasmodesma for their benefits while keeping the cell membrane intact to prevent the initiation of plant defense responses. These tactics are well employed by plant viruses (non-living entities and entirely dependent on the host’s living tissues).
Plant Viruses Interactions with Plasmodesmata
Plant viruses are obligatory parasites that completely depend on their host living cells for their multiplication. Once they get entry into their host plant through vectors (insects), their main purpose is to infect the host and utilize its metabolism for their own purpose. Plant viruses infect plants from moving one cell to another. For their movement from an infected cell to adjacent healthy cells, viruses must encode a specialized protein referred to as “movement proteins” (MV) to cross the microchannel.
Numerous studies on Tobacco mosaic virus (TMV), Potato virus X (PVX), and Cucumber mosaic virus (CMV) suggested that viruses use non-structural movement proteins that interact with plasmodesmata (PD)and endoplasmic reticulum (ER)for transfer to nearby healthy cells.
Viruses movement proteins (MP)are diverse in nature and perform the same functions:
- Encoded by the virus and interact with the virus genome for its transport
- Interact with plasmodesmata (PD) especially with the endoplasmic reticulum
- Target the function of PD and modify its molecule trafficking ability and make its pore more favorable for virus movement to neighboring cells. This process is called “gating”.
Gating
Plant pathogens interact with plasmodesmata (PD) and enhance the small molecule trafficking and diffusible proteins for their own benefits and this phenomenon is called gating. Despite being many studies on viruses (TMV, PVX, CMV) interactions with plasmodesmata, very little information is present about “How do plant viruses gate?”
Different studies suggest that pathogens use gating for infection and long-distance spread. Few MPs of viruses are able to gate PD and while other movement proteins have zero roles in gating. Various models of gating have been proposed by plant virologists. One is proposed by (Epel, 2009). According to this model, plant viruses gate PD by altering the structure of plasmodesmata pores. This alteration is facilitated by the degradation of callose: it is a plant polysaccharide and acts as a temporary cell wall under unfavorable conditions. Callose degradation enhances the size of PD pores and supports the virus molecule's trafficking.
Keynotes:
Plant viruses move from cell to cell by interacting with cellular mechanisms and these mechanisms control the plasmodesmata pore size limit.
References: